Synaptic Connectivity
Background: The long term goals of the lab are to understand the molecules and developmental programs that regulate neuronal development and wiring. In a previous study, we investigated the biology of the novel interactions between two subfamilies of the immunoglobulin superfamily in Drosophila melanogaster (in collaboration with Christopher Garcia at Stanford and Engin Ozkan at the University of Chicago; Ozkan et al., 2013): the defective proboscis extension response proteins (Dprs), encompassing 21 members, and the 9-member Dpr-interacting proteins (DIPs). These proteins are expressed in unique subsets of neurons throughout the nervous system and are implicated in various developmental processes. Previously, we found that an interacting Dpr-DIP pair functions at various developmental stages including motor neuron development at the larval neuromuscular junction (NMJ) and wiring and cell survival in the pupal optic lobe (Carrillo et al., 2015).

Visual circuits
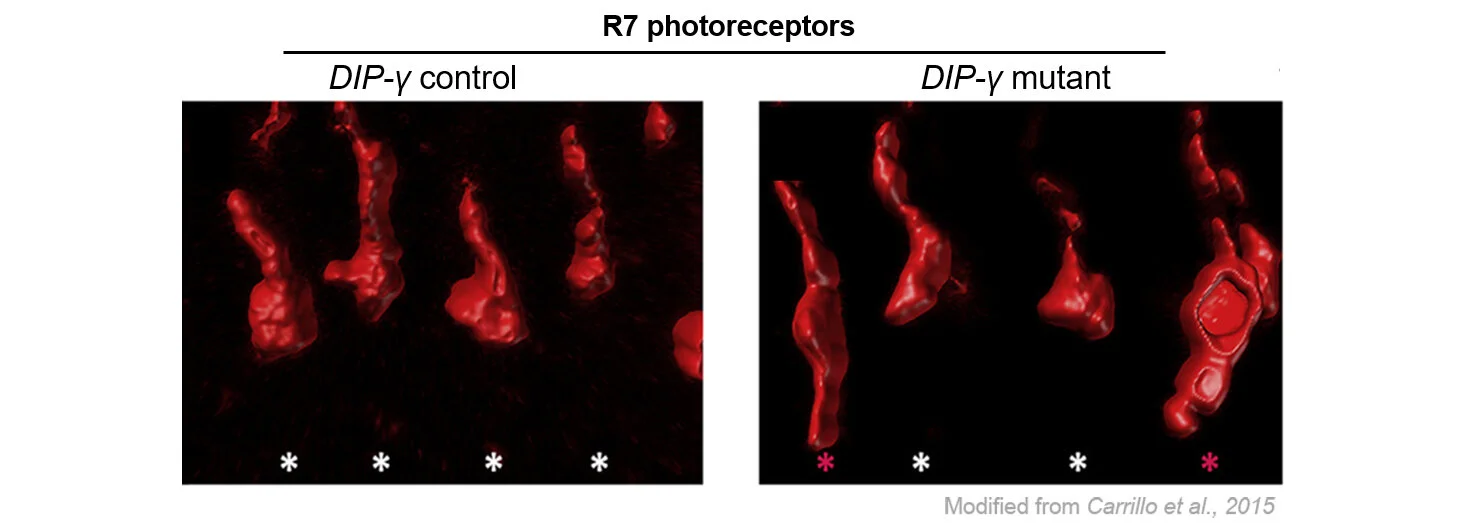
Proper neuronal connectivity is necessary for brain function. As part of the Drosophila visual system, photoreceptors receive sensory information and relay this signal to downstream neurons in order to respond accordingly. In a prior study, we found that cell surface proteins regulate synaptic connectivity of a subset of photoreceptors to a target neuron: loss of dpr11 and DIP-γ, interacting cell surface proteins, results in yellow R7 photoreceptors axons overshooting their Dm8 synaptic partners (shown by red asterick in the figure). Our lab and others reported that many synaptic partners express cognate Dpr-DIP pairs, and we are exploring the generality of these mechanisms to assemble visual circuits.
Neuromuscular circuits
Similar to complex brain circuits, the Drosophila larval neuromuscular circuit encounters connectivity challenges. In this circuit, developmental programs must instruct 35 motor neurons in the embryonic ventral nerve cord to extend their axonal projections to the periphery where they must decide which of the 30 muscle fibers to innervate. Despite over 40 years of investigations, we still lack a complete understanding of the mechanisms that underlie neuromuscular circuit assembly. In a recent study, we identified Dpr10 (in muscles) as one of the long-sought targeting cues that direct recognition of specific muscle as targets, while DIP-α is the corresponding receptor on the motor neurons that innervate these muscles. We showed that Dpr10-DIPα interaction is crucial for motor neuron innervation on muscle 4 (figure shows loss of 1s innervation on muscle 4) (Ashley et al., 2019). We are currently screening additional Dprs and DIPs to systematically analyze how these cell adhesion molecules establish synaptic connective to drive larval locomotion.

Ventral nerve cord
While motor neurons send their axons to muscles, they receive information on their dendrites from interneurons in the ventral nerve cord. Extensive electron microscopy reconstruction has defined connectivity maps upstream of motor neurons allowing us to examine synaptic recognition molecules that wire these circuits. Based on the unique expression patterns of Dprs and DIPs, we will combine loss-of-function studies with stochastic labeling (figure shows single cell labeling in VNC) to uncover Dpr-DIP interactors that instruct interneuron-motor neuron synaptic recogniton.
